Grupos sexuales activos en vacas posparto de ganadería mestiza de doble propósito
Lílido N. Ramírez-Iglesia1, Freddy B. Viera Rosales1, Jesús A. Martínez2, Adelina Díaz de Ramírez1 José G. Morillo Luque1, Rafael Román3 y Eleazar Soto-Belloso1
1 Universidad de Los Andes. Apartado Postal 198
Trujillo 3150, estado Trujillo. Venezuela
2 Ejercicio Privado
3 Facultad de Ciencias Veterinarias
La Universidad del Zulia
Maracaibo, estado Zulia. Venezuela
Recibido y aceptado dentro del contexto del
XIII congreso de Producción e Industria Animal 2006
RESUMEN
En la ganadería bovina mestiza de doble propósito (GDP), la deficiente detección de celos afecta la difusión de la inseminación artificial y la eficiencia reproductora. Para estudiar la formación y características de los grupos sexuales activos (GSA), en una finca ubicada en una zona de bosque húmedo tropical de la cuenca del Lago de Maracaibo, se estudió un rebaño de 300 vacas posparto Brahmán Rojo (Bos indicus) x Holstein Rojo (Bos taurus), ordeñadas manualmente con apoyo del becerro dos veces al día, los cuales pastorearon potreros de pasto Alemán (Echinocloa polistachia) y Guinea (Panicum maximun), acompañadas por toros receladores 25:1. Las vacas fueron observadas cuatro veces de 5:00 a 6:00 AM y de 2:00 a 6:00 PM en los corrales y de 6:00 a 10:00 AM y de 6:00 a 7:00 PM en los potreros. Se consideró GSA, el constituido por no menos de tres animales en interacción de monta de celo. Se detectaron 51 GSA, resultando el 76% heterosexual y 24% homosexual, 69% se observaron en corrales y 31% en potreros, 41% en la mañana y 59% en la tarde. Los GSA acumularon el 67% de las montas de celo y la duración media de esos GSA fue de 20 ± 2,8 minutos. La cadena de montas en la mayoría de los grupos heterosexuales fue iniciada con la monta de un toro recelador. La detección, registro y evaluación de los GSA en la GDP puede aportar a mejorar la eficiencia de la detección del celo.
Palabras clave: grupos sexuales activos, detección celo, vacas doble propósito, posparto, clima tropical.
Sexually active groups in dual purpose postpartum crossbred cattle
Summary
In dual purpose crossbreed cattle (DPC), deficient estrus detection affects the artificial insemination diffusion and reproductive efficiency. To study the formation and characteristics of sexually active groups (SAGs), 300 crossbred postpartum Red Brahman (Bos indicus) x Red Holstein (Bos taurus) cows were studied and milked manually twice a day with support from the calf, grazing German (Echinocloa polistachia, 12% crude protein) and Paez (Brachiaria mutica) grass in pasture lands, accompanied by teaser bulls at a 25:1 ratio. They were observed four times, from 5am to 6:00 am at cement pen area, and from 6:00 am to 10:00 am in the pasture area. Finally from 2:00 pm to 6:00 pm, and from 6:00 pm to 7:00 pm at the cement pen and pasture area respectively. This was performed in a commercial farm located in a Humid Tropical Forest area in the Maracaibo Lake basin, in the Bolivarian Republican of Venezuela. Only the groups constituted of no less than three animals in standing mount interaction were considered SAGs. 51 SAGs were detected, 76% heterosexual, and 24% homosexual, 69% were observed in cement pens and 31% at pasture area, 41% in the morning and 59% in the afternoon. SAGs accounted for 67% of the standing estrus observed; the duration time of those SAGs was of 20±2, 8 minutes. In most of the heterosexual SAGs the teaser bull was beginning the mounting chain. The detection, registration and evaluation of SAGs in DPC can contribute to improve the efficiency of estrus detection.
Key words: sexually active group, oestrus detection, dual purpose crossbred cows, postpartum, tropical climate.
INTRODUCCIÓN
En la ganadería mestiza de doble propósito (GDP), el ciclo estral de las vacas tiene una duración de 18 a 29 días, el celo muestra una media de 16,4 ± 5,2 horas y la ovulación ocurre entre 25 y 30 horas después de iniciado el estro, manteniendo estos eventos fisiológicos una duración variable según la época del año y el genotipo del animal (González-Stagnaro, 1992; Domínguez et al., 2004).
En la GDP, el anestro posparto y la deficiente detección de celos son los principales factores que afectan y limitan la difusión y el éxito de la técnica de inseminación artificial (IA), los cuales combinados con las bajas tasas de fertilidad, constituyen la trilogía esencial que debe ser resuelta para el mejoramiento de la eficiencia reproductora de estos rebaños tropicales (GonzálezStagnaro, 1992; 2002; Soto-Belloso et al., 2000; 2002). Igualmente a los errores y deficiencias en la detección del celo, también se han responsabilizado de ser una causa importante de problemas reproductivos y subfertilidad en los rebaños mestizos (González-Stagnaro, 2002) y bajas tasas de servicios en IA (Bo y Baruselli, 2002). Ante estas circunstancias y para evitar la escasa eficiencia en la detección de celos, se han desarrollado una serie de protocolos hormonales destinados a inducir o sincronizar el celo y la ovulación, favoreciendo la IA sistemática a tiempo fijo, sin necesidad de detectar los celos (Bo y Baruselli, 2002). Sin embargo, se han reportado mejores tasas de concepción en las vacas inseminadas luego de la detección visual del celo que en aquellas inseminadas a ciegas, tanto en ganaderías de leche (DeJarnette et al., 2001) como de carne (Bo y Baruselli, 2002). Adicionalmente, se han encontrado altas y significativas correlaciones entre el nivel de estrógenos y los signos del celo (Lyimo et al., 2000) y mejores tasas de preñez (Rae et al., 1999) en la observación visual cuando se compara con otros métodos de detección.
En la GDP, considerada de baja inversión, sostenible y ecológica, se ha consolidado la técnica de la observación visual de la monta como la más apta para la detección del celo y, sobre ello, se apoyan los programas de IA para el mejoramiento genético del rebaño. Sin embargo, en estos sistemas de manejo, la cuantificación de los niveles de progesterona en plasma y leche solo han mostrado una detección correcta del celo en las vacas entre 83,3 y 95,5% (Soto-Belloso et al., 2000), ratificando el significado de la deficiente detección del celo como el mayor problema de los sistemas de GDP mejorados y la importancia de utilizar conceptos del comportamiento que permitan alcanzar mejores tasas de detección (González-Stagnaro, 2002).
La cultura y práctica ganadera en los ambientes tropicales, al igual que en los templados, ha llevado a los productores a confiar, fundamentalmente, en la inmovilidad de la vaca en celo al ser montada por un toro o por una de sus compañeras del rebaño, restando importancia a los otros signos de conducta sexual que expresan las hembras bovinas durante este estado fisiológico único y fundamental para reproducirse (Sepúlveda et al., 2002). Tanto en el estro natural como en el inducido hormonalmente se ha detectado que la presencia en el rebaño de más de una hembra en celo desencadena una interacción sexual de montas y otras conductas estimulantes del reclutamiento e intervención sexual de otras vacas, con la consiguiente formación transitoria y móvil de grupos homosexuales o heterosexuales conocidos como grupos sexuales activos (Galina et al., 1996; Castellanos et al., 1997; Orihuela, 2000, Gonzáles-Stagnaro, 2000). Se señalan como mínimo agrupamiento el integrado por dos animales (Castellanos et al., 1997; González-Stagnaro, 2000).
A pesar de la importancia de la detección visual del celo en los sistemas mejorados de GDP, la conducta sexual de esos bovinos mestizos ha sido objeto de pocos estudios conocidos y publicados. Un reporte sobre los principales y más frecuentes signos del celo observados en el ganado mestizo de doble propósito fue publicada por Ramírez-Iglesia et al. (2002). Dado el papel que la observación visual de la aceptación inmóvil de la monta desempeña en la identificación de la hembra en estro, este trabajo tuvo como objetivo el estudio de la formación, composición, lugar y características de los grupos sexuales activos durante la detección del primer celo espontáneo posparto, bajo las condiciones rutinarias de manejo de un rebaño mestizo con ordeño manual y la aplicación de un programa de inseminación artificial.
MATERIALES Y MÉTODOS
Se utilizó un rebaño comercial de 300 vacas mestizas de doble propósito con uno a ocho partos y predominancia racial de animales provenientes del cruce alterno entre Brahman rojo (Bos indicus) X Holstein Rojo (Bos taurus). La finca estaba ubicada en una zona de bosque húmedo tropical de la cuenca del Lago de Maracaibo, situada a unos 9o 27 N y 72o 20 O con precipitaciones de 1.800 mm por año, temperatura media anual de 29ºC y una humedad relativa del 50 a 70%. Las hembras estaban distribuidas en dos lotes de 150 animales cada uno y fueron ordeñadas manualmente con apoyo del becerro dos veces al día, de 3:00 a 6:00 AM y de 3:00 a 6:00 PM. Presentaron una condición corporal al primer celo posparto de 3,2 ± 0,2 en una escala 0 = emaciada a 5 = obesa, según el tejido adiposo en la zona caudo-pélvica (Mulvany, 1981). Las vacas estaban acompañadas permanentemente por toros mestizos receladores en una proporción de 25:1, sometidas a un programa de inseminación artificial con la regla AM-PM, con registros de los principales eventos reproductivos y productivos, revisión posparto a los 30 días y cada mes para el diagnóstico de gestación y evaluación reproductiva. Se alimentaron a pastoreo en praderas de pasto alemán (Echinocloa polistachia) con un 12% de proteína cruda, paja páez (Brachiaria mutica) y pasto estrella (Cynodon lemfuensis) y también se les suministró sal yodada, minerales y agua ad libitum. Los corrales y sala de ordeño tenían piso rústico de cemento. Las hembras diagnosticadas gestantes eran trasladadas a otro rebaño de ordeño de vacas gestantes.
Para detectar la aceptación quieta de la monta como indicador el primer celo posparto natural (PCP) y la formación de los grupos sexuales activos, se observaron diariamente durante diez horas discontinuas, cinco en la mañana y cinco en la tarde de acuerdo a los siguientes períodos o rutinas: en los corrales, de 5:00 a 6:00 AM y de 2:00 a 6:00 PM y en los potreros, de 6:00 a 10:00 AM y de 6:00 a 7:00 PM. También, se registraron las montas detectadas en las callejuelas durante el traslado de los animales. Las vacas estaban identificadas con un número visible marcado a fuego en el lomo del lado derecho y tatuaje en la oreja izquierda. Las observaciones se realizaron durante 25 días entre el 26 de junio y el 23 de agosto, periodo correspondiente a la época seca o de verano.
Se consideró "vaca en celo" a aquella que aceptó inmóvil la monta de una de sus compañeras o de un toro recelador (Hurnik et al., 1995). Se consideró como "grupo sexual activo" (GSA), el constituido por no menos de tres animales que interactuaron con montas y aceptación de la monta durante alguno de los cuatro periodos de observación, definición basada en los reportes de diversos autores (Galina et al., 1996; Castellanos et al., 1997; Orihuela, 2000, Gonzáles-Stagnaro, 2000). Dado que se ha señalado como grupo sexual activo el constituido por dos o más animales (Castellanos et al., 1997; González-Stagnaro, 2000), en este trabajo se diferencian los grupos formados por dos individuos de aquellos con mayor número de integrantes. Bajo este criterio, se consideró como "pareja sexual activa" (PSA) a aquella con participación de solo dos animales. La intensidad de la actividad sexual se estableció mediante la "intensidad de monta" (IM) en el grupo, indicador que relaciona el total de montas con el número de animales integrantes del mismo (total de montas del grupo /numero de individuos del grupo). La duración del GSA se estimó por el intervalo de tiempo transcurrido entre la primera y la última monta de celo en el grupo. La duración de la PSA no se determinó.
Se registró el día, hora y lugar de la monta, el número de identificación y sexo del animal que montó, número de la vaca receptora de la monta, con lo cual, se determinó el número de animales comprometidos en la interacción sexual del GSA, el lugar de formación, la característica sexual, el número de montas de celo, número de montas aceptadas por cada vaca y el tiempo de duración del grupo.
Los datos fueron analizados con el paquete estadístico SAS (SAS, 1989) usando el procedimiento FREQ con prueba del chi-cuadrado. Para ello, se clasificaron de acuerdo al sexo de sus integrantes en homosexuales y heterosexuales, por el lugar de formación del grupo en corrales y potreros, por la hora del día en mañana y tarde, por el número de animales comprometidos en la interacción sexual de la monta de celo en GSA y PSA. Los datos fueron procesados en el Centro de Computación de la Universidad de los Andes.
RESULTADOS Y DISCUSIÓN
En el Cuadro 1 se describe la distribución de los agrupamientos sexuales activos formados de acuerdo a la clasificación establecida, donde se observa la conformación de un 60% (76/127) de PSA y 40% (51/127) de GSA. De estos últimos se destaca que un 31% (39/127) fueron de tipo heterosexual, 28% (35/127) se detectaron en los corrales y el 24% (30/127) en horas de la tarde; en tanto que, para las PSA los porcentajes fueron del 30% (38/127), 41% (52/127) y 40% (51/127) para las categorías heterosexual, corrales y horas de la tarde respectivamente.
|
Cuadro 1. Descripción de las características de los agrupamientos sexuales activos posparto en ganado mestizo de doble propósito |
||||||
| Características |
PSA |
GSA |
Total |
|||
|
n |
% |
n |
% |
n |
% | |
| Tipo interacción | ||||||
|
Heterosexual |
38 |
30 |
39 |
31 |
77 |
61 |
|
Homosexual |
38 |
30 |
12 |
9 |
50 |
39 |
|
Total |
76 |
60 |
51 |
40* |
127 |
100 |
| Lugar | ||||||
|
Corrales |
52 |
41 |
35 |
28 |
87 |
69 |
|
Potreros |
24 |
19 |
16 |
12 |
40 |
31 |
|
Total |
76 |
60 |
51 |
40 |
127 |
100 |
| Hora | ||||||
|
Mañana |
25 |
20 |
21 |
16 |
46 |
36 |
|
Tarde |
51 |
40 |
30 |
24 |
81 |
64 |
|
Total |
76 |
60 |
51 |
40 |
127 |
100 |
|
Montas (n) |
85 |
33 |
176 |
67 |
261 |
100 |
|
Duración grupo min (X±Ee) |
ND |
20±2,8 |
ND |
|||
|
Intensidad de Monta (X±Ee) |
0,56 ± 0,02 |
0,84 ± 0,03 |
0,67 ± 0,02 |
|||
|
PSA: Pareja sexual activa constituida por
dos animales. GSA: Grupo sexual activo tres y más animales.
ND= No determinado.
*= P<0.01. Min= minutos. Intensidad de la monta=número de montas en el grupo/número de animales en el grupo. |
||||||
La intensidad de monta (IM) fue de 0,56 ± 0, 02 y 0,84 ± 0,03 montas/grupo para PSA y GSA respectivamente, lo cual, es explicable por que en la PSA participaron solo dos animales que exhibieron una (88%) y dos montas (12%). La variable GSA tomó valores extremos de 0,5 a 1,5 montas/grupo, lo que sugiere una mayor actividad de monta en ese grupo; estando integrado el 92% de esos GSA por tres, cuatro y cinco animales, que representaron el 49%, 20% y 23%, respectivamente, mientras que ese porcentaje fue de solo 8% para aquellos con mas de cinco individuos. Estos tamaños de grupos coinciden con los reportados luego del estudio de la conducta sexual durante el estro de un rebaño experimental de vacas de la raza cebú (Castellanos et al., 1997), quienes señalaron que el mayor número de grupos sexuales identificados se formaron con pocos miembros (3,5 ± 1,2 animales). En este estudio se detectaron entre dos y nueve montas por grupo, con un total del 82% para aquellos grupos con dos, tres y cuatro montas de estro y valores de 39, 20 y 23%, respectivamente. En el Cuadro 1 se observa que de las 261 montas anotadas durante este estudio, 176 (67%) de ellas correspondieron a los GSA y 85 (33%) a las PSA. Estos hallazgos son explicables debido a que la presencia al mismo tiempo de dos o más vacas en celo estimuló la actividad de monta, tal como ha sido reportado para ganaderías especializadas de leche (Pennington et al., 1985) y de carne (Galina et al., 1996). Naturalmente, el mayor número de hembras en estro crea condiciones favorables para la formación de los GSA y la cadena de montas que se desatan con el reclutamiento o estimulación de otras vacas próximas o en celo.
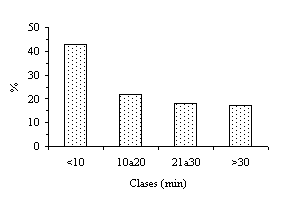 |
| Figura 1. Duración de grupos sexuales activos envacas posparto de ganadería mestiza de doble propósito. |
La duración (media ± error estándar) de actividad en los GSA fue de 20 ± 2,8 minutos. En la Figura 1 se presenta una descripción de ella, observándose un 43 y 22% para las clases =10 y entre diez y veinte minutos, acumulándose una frecuencia de 65% para aquellos GSA con una duración menor de 20 minutos. Aunque no se tiene una explicación de la corta duración de estos agrupamientos sexuales activos, ella puede estar relacionada con el número de animales de animales en estro, la intensidad y la fase del celo (Hurnik et al., 1975; Alexander et al., 1984; Hurnik y King, 1987). Esa relativa breve duración de los GSA constituye una interesante información a tener en cuenta para mejorar la eficiencia de la detección del celo.
Se observó que 41% de los GSA se formaron en la mañana y 59% en la tarde. Para las PSA, esos guarismos fueron de 33 y 67% en la mañana y tarde, respectivamente (Cuadro 2). Estos porcentajes fueron ligeramente superiores al 32 y 33% de los celos matutinos reportados por González-Stagnaro (1992) y por Galina et al. (1982), respectivamente. Recientemente Góngora y Hernández (2006) comunicaron un 72% de los estros en horas de la mañana. Estos resultados son muy superiores a los observados en el presente estudio. Los citados autores, estudiando experimentalmente el comportamiento estral en novillas sincronizadas de las razas cebú Brahman y Criollas San Martineras, mantenidas a potrero y observadas continuamente entre las 5:00 AM y 7:00 PM, registraron el 72% de los estros entre las 5:00 AM y las 11:00 AM, en desacuerdo con lo observado en este estudio, atribuible a que estas eran vacas sometidas al manejo rutinario del ordeño. Por otro lado, destaca la importancia de la detección del 31% de estos grupos sexuales a nivel de los potreros, donde rutinariamente no son observados, lo que sugiere la necesidad de instituir esta práctica en forma habitual para mejorar la eficiencia en la detección del celo.
|
Cuadro 2. Frecuencias de los GSA y las PSA de acuerdo al tipo de interacción sexual, lugar y hora de detección de la monta de celo |
|||||
| Tipo de Interacción |
Lugar de detección |
Total | |||
|
Corrales |
Potreros |
||||
|
Mañana |
Tarde |
Mañana |
Tarde | ||
|
% |
|||||
|
Grupos Sexuales Activos (GSA) |
|||||
|
Heterosexual |
6 |
47 |
23 |
0 |
76 |
|
Homosexual |
4 |
12 |
8 |
0 |
24 |
|
Total |
10 |
59 |
31 |
0 |
100 |
|
Parejas Sexuales Activas (PSA) |
|||||
|
Heterosexual |
8 |
29 |
10 |
3 |
50 |
|
Homosexual |
3 |
29 |
12 |
6 |
50 |
|
Total |
11 |
58 |
22 |
9 |
100 |
En el citado Cuadro 2, se destaca la detección solo en horas de la mañana de los GSA conformados a nivel de potreros y la mayor formación, tanto de los GSA como de las PSA en los corrales durante las horas de la tarde. Este último hallazgo puede ser atribuible al confinamiento transitorio, que pudo ser capaz de producir la concentración de feromonas sexuales y otros olores que por si mismas o en combinación con señales acústicas, visuales y de contacto físico pueden estimular la conducta sexual (Rekwot et al., 2001) y de monta (González-Stagnaro, 2000), creándose condiciones favorables para la formación de los GSA, a lo cual, puede contribuir el mayor número de horas de observación en este lugar donde los animales fueron alojados antes de ingresar a la sala de ordeño. Si bien, en estos climas tropicales y para este tipo de animal mestizo, no se ha definido claramente la zona confort de temperatura ambiente, en el ganado lechero se ha reportado un incremento curvilíneo de la actividad de monta a medida que se eleva la temperatura diaria dentro de la zona confort para eso grupos raciales (Gwazdauskas et al., 1983). Estas relaciones genotipo-ambiente-conducta sexual estral deben ser indagadas para la ganadería mestiza de doble propósito.
La mayor frecuencia de GSA heterosexuales formados en los potreros en horas de la mañana, además de ser las más frescas del día, lo que sin duda es favorable para este tipo de actividad sexual, pudo estar asociado con un mayor campo visual del toro recelador, quien puede usar la visión de la actividad sexual de aceptación de la monta de las hembras, como principal señal, para identificarlas cuando se encuentra en celo (Geary y Reeves, 1992) e integrarse al GSA.
El 41% de los GSA heterosexuales se conformaron con un toro y dos, tres y cuatro vacas que representaron el 26, 10 y 5%, respectivamente. El 56% tuvieron una composición de dos toros y una, dos, tres, cuatro y seis vacas, significando el 23, 13, 13, 5 y 2%, respectivamente. Solo el 3% de estos agrupamientos tuvieron una composición de tres toros y dos vacas, en tanto que, para las PSA el 50% fueron heterosexuales. En la composición de los GSA homosexuales participaron tres, cuatro, cinco y ocho vacas, correspondiendo un porcentaje del 58,4 8,3 25 y 8,3%, respectivamente. En las PSA, el 50% de ellas fueron homosexuales.
En el Cuadro 3 se presenta el porcentaje, el tipo o característica sexual del grupo, la secuencia cronológica de observación de la monta de celo, el número de montas en el grupo, el tiempo de duración de algunos de esos agrupamientos sexuales activos y el número de montas aceptadas por la vaca. Se registró que la mayoría de los grupos heterosexuales se conformaron con uno y dos toros, mientras que los homosexuales con tres a cinco vacas. Igualmente se observó que la mayoría de los GSA heterosexuales se iniciaron con la monta de un toro recelador a una vaca en celo. En la secuencia de montas se aprecia la presencia de vacas receptoras-montadoras y solamente receptoras. En la última columna del Cuadro 3 se observa el registro del número de montas para cada hembra del grupo, detectándose la presencia de vacas montadoras, más no recíprocamente receptoras de la monta (0), por lo cual, esas hembras montadoras no fueron consideradas en celo ni sujetos de la IA.
Cuadro 3. Tipo de interacción sexual y característica de la monta de celo en grupos sexuales activos de vacas mestizas posparto de doble propósito.
La detección en este estudio de vacas exclusivamente montadoras, exclusivamente receptoras y receptoras-montadoras es un fenómeno conductual que debe ser objeto de mayores estudios en la GDP para relacionar esa conducta estral con el tamaño y el estatus fisiológico del folículo y del eje hipotálamo-hipófisis-ovario, tal como ha sido sugerido por Maquivar et al. (2002), quienes reportaron un 50% de vacas montadoras y un 50% de vacas montadoras-receptoras de monta con folículos ováricos entre 4,9 y 11,2 mm, insinuando un desacuerdo entre el tamaño de los folículos, la secreción de estrógenos y su uso como indicadores de la conducta estral de las vacas.
No se descarta en estos agrupamientos sexuales activos posparto, la presencia de conductas alelomiméticas o imitativas (Hurnick et al., 1995; Pires et al., 2003) y otros factores de comportamiento de los bovinos, como los inherentes a la existencia de un orden y jerarquías sociales. De igual manera, aquellas relacionadas con el tamaño y composición del rebaño (Galina et al., 1996; Solano et al., 2005), interacciones de facilitación o integración social, de estimulación social, sinergismo conductual, preferencias sexuales individuales, selectividad y otras relacionadas con el manejo, la edad del rebaño en ordeño, nutricionales, ambientales y genéticas que afectan la manifestación del estro, al igual que la eficiencia y precisión de su detección (Orihuela, 2000).
La formación de los GSA es un fenómeno fisiológico conductual de gran importancia para una GDP en cuyos sistemas mejorados con IA, el signo confiable, patognomónico y único evidente para que la vaca sea inseminada es la inmovilidad de la vaca al ser montada por un toro o por una de sus compañeras.
CONCLUSIONES y RECOMENDACIONES
La formación de grupos sexuales activos es un fenómeno de la conducta sexual de los bovinos, presente en la ganadería mestiza de doble propósito. Mayoritariamente se conformaron con tres a cinco animales, quienes manifestaron una gran actividad de montas de celo. La mayoría tuvo una duración menor de 20 minutos y fueron fácilmente detectados tanto a nivel de los corrales como en los potreros.
La observación visual requiere ajustar los períodos de detección de celo basándose en el ritmo impuesto por la rutina del ordeño, de acuerdo a las horas de luz del día, la observación rutinaria en los potreros y el diseño de planillas especiales de registros con evaluación diaria de la actividad de las vacas receptoras, receptoras-montadoras y exclusivamente montadoras.
Se requieren mayores estudios sobre la calidad del celo, la consecuente ovulación, las conductas alelomiméticas, el orden y jerarquías sociales y otros fenómenos fisiológicos, conductuales y relaciones ambientales que puedan afectar la eficiente y correcta detección de la vaca mestiza en celo.
AGRADECIMIENTOS
Al CDCHT-ULA por el financiamiento a través del proyecto NURR-C-304-01-03-F y a la Agropecuaria Santa Ana, propietaria de la finca Mompox por el apoyo brindado.
LITERATURA CITADA
Alexander T. J., P. L. Senger, J. L. Rosenberger y D. R. Hagen. 1984. The influence of the stage of the estrous cycle and novel cows upon mounting activity of dairy cattle. J. Anim. Sci., 59(6): 1430-1439
Bo G. A. y P. S. Baruselli. 2002. Programas de inseminación artificial a tiempo fijo en el ganado bovino en regiones subtropicales y tropicales. En González-Stagnaro C., E. Soto Belloso y L. Ramírez-Iglesia (Eds). Avances en la Ganadería de Doble Propósito. Fundación GIRARZ. Ediciones Astro Data. Maracaibo, Venezuela. pp. 497-514.
Castellanos F., C. S. Galina, J. A. Orihuela, R. Navarro-Fierro y R. Mondragón. 1997. Estrous expression in dairy cows and heifers (Bos taurus) following repeated PGF2a injection and choice of selecting a mounting partner. App. Anim. Beh. Sci., 51: 29-37.
DeJarnette J. M., R. R. Salverson y C. E. Marshall. 2001. Incidence of premature estrus in lactating dairy cows and conception rates to standing estrus or fixed-time inseminations after synchronization using GnRH and PGF2a. Anim. Rep. Sci., 67: 27-35.
Edmonson A. J., I. J. Lean, L. D. Weaver, T. Farver y G. Webster. 1989. A body condition scoring chart for Holstein Dairy Cows. J. Dairy Sci., 72:68-78.
Domínguez C., N. Martínez y O. Colmenares. 2004. Características reproductivas de rebaños bovinos de doble propósito en los llanos centrales. Zootecnia Trop., 22(2): 133-145.
Galina C. S., A. Calderón y M. McClosky. 1982. Detection of signs of estrus in the Charolais cow and its Brahman cross under continuous observation. Theriogenology, 17(5): 485-498.
Galina C. S., A. Orihuela e I. Rubio 1996. Behavioural trends affecting oestrus detection in Zebu cattle. Anim. Rep. Sci., 42: 465-470.
Geary T. W. y J. J. Reeves. 1992. Relative importance of vision and olfaction for detection of estrus by bulls. J. Anim. Sci., 70: 2726-2731.
Góngora A. y A. Hernández. 2006. Comportamiento sexual, duración del estro y del ciclo estral en novillas criollas sanmartineras y brahman del piedemonte llanero colombiano. Livestock Res. Rural Dev., 18(1) http://www.cipav.org.co/lrrd/lrrd18/1/gong18001.htm
González-Stagnaro C. 1992. Fisiología reproductiva en vacas mestizas de doble propósito. En González-Stagnaro C. (Ed). Ganadería Mestiza de Doble Propósito. Fundación GIRARZ. La Universidad del Zulia. Ediciones Astro Data. Maracaibo, Venezuela. pp. 153-188.
González-Stagnaro C. 2000. Utilice el grupo sexual activo (GAS) y mejore la detección de los celos en las vacas. Venezuela Bovina, 15(47): 42-45.
González-Stagnaro C., N. Madrid-Bury y J. Goicochea-Llaque. 2002. Sistemas de manejo y eficiencia en la detección de celos en rebaños doble propósito. Rev. Cien. Fac. Cien.Vet. LUZ, 12(Supl. 2): 455-463.
Gwazdauskas F. C., J. A. Lineweaver y M. L. McGilliard. 1983. Environmental and management factors affecting estrous activity in dairy cattle. J. Dairy Sci., 66: 1510-1514.
Hurnik J. F., G. J. King y H. A. Robertson. 1975. Oestrus and related behaviour in postpartum Holstein cows. Appl. Anim. Ethol,, 2: 55-58.
Hurnick J. F. y G. J. King. 1987. Estrous behavior in confined beef cows. J. Anim. Sci., 65: 431-438.
Hurnik J .F., A. B. Webster y P.B. Siegel. 1995. Dictionary of Farm Animal Behavior. 2da ed. Iowa State University Press.
Lyimo Z. C., M. Nielen, W. Ouweltjes, T. A. M. Kruip y C. M. Van Eerdenburg. 2000. Relationship among estradiol, cortisol and intensity of estrous behavior in dairy cattle. Theriogenology, 53: 1783-1795.
Maquivar M., C. S. Galina y A. Orihuela. 2002. Cows treated with synchromate B may cluster their sexual behavior independent of follicular growth at time of oestrus. Physiol. Behavior, 76: 199-203.
Mulvany P. 1981. Dairy cow condition scoring. Nat. Inst. Res. Dairying. Handout No. 4468. Shinfield, Reading, UK.
Orihuela A. 2000. Some factors affecting the behavioral manifestation of oestrus in cattle: A review. Appl. Anim. Behaviour Sci., 70: 1-16.
Pennington J. A., J. L. Albright y M. A. Diekman. 1985. Sexual activity of Holstein cows: Seasonal effects. J. Dairy Sci., 68: 3023-3030.
Pires M., F. Ávila, N. G. Alves, J. M. Silva Filho, L. S. A. Camargo y R.S. Verneque. 2003. Estrus behaviour in Gir cows (Bos taurus indicus). Arq. Bras. Med. Vet. Zootec., 55(2): 187-196.
Rae D. O., P. J. Chenoweth, M. A. Giangreco, P. W. Dixon y F. L. Bennett. 1999. Assessment of estrus detection by visual observation and electronic detection methods and characterization of factors associated with estrus and pregnancy in beef heifers. Theriogenology, 51: 1121-1132.
Ramírez-Iglesia, L. N., F. B. Viera-Rosales, J. A. Martínez, A. Díaz de Ramírez, E. Soto-Belloso. 2002. Conducta sexual y signos del celo en ganadería mestiza de doble propósito. Rev. Cien. Fac. Cien. Vet. LUZ, 12(Supl. 2): 431-433.
Rekwot P. I., D Ogwu, E. O. Oyedipe y V. O. Sekoni. 2001. The role of pheromones and biostimulation in animal reproduction. Anim. Rep. Sci., 65: 157-170.
SAS Institute. 1989. SAS/STAT Users Guide. Versión 6. 4ta ed, SAS Institute Inc. Cary, NC.
Sepúlveda Becker N. G. y E. Rodero Serrano. 2002. Evaluación de la detección del celo en explotaciones lecheras. Rev. Cien. Fac. Cien. Vet. LUZ, 12(3): 169-174.
Solano J., A. Orihuela, C. S. Galina, F. Montiel y F. Galindo. 2005. Relationship between social behaviour and mounting activity of Zebu cattle (Bos indicus). Appl. Anim. Behaviour Sci., 94: 197203.
Soto-Belloso E., G. Portillo Martínez, A. De Ondiz, N. Rojas, G. Soto Castillo, L. Ramírez Iglesia y F. Perea Ganchou. 2002. Improvement of reproductive performance in crossbred zebu anoestrus primiparous cows by treatment with norgestomet implants or 96 h calf removal. Theriogenology, 57: 1503-1510.
Soto-Belloso E., G. Portillo Martínez, A. De Ondiz Sánchez, N. Rojas, G. Soto Castillo, L. N. Ramírez-Iglesia, J. Aranguren y F. Perea Ganchou. 2000. Evaluación del comportamiento reproductivo mediante el uso de la progesterona por radioinmunoanálisis en vacas mestizas Cebú bajo programas de inseminación artificial en Venezuela. Rev. Cien. Fac Cien. Vet. LUZ, 10(5): 391-398.