Efecto de la suplementación con Saccharomyces cerevisiae sobre el comportamiento reproductivo en rebaños lecheros de la zona alta del estado Mérida
José Rivas1, Pedro Bastidas2, Thais Díaz2, Martín Hahn1 y Alexis Moya3
1 Departamento de Producción Animal, Facultad de Ciencias Veterinarias
Universidad Central de Venezuela. Maracay, Aragua. Venezuela
2 Instituto de Reproducción Animal e Inseminación Artificial Facultad de
Ciencias Veterinarias, Universidad Central de Venezuela, Maracay, Aragua 2101
3 MoyaMix C.A.
RESUMEN
El objetivo del presente estudio fue determinar el efecto de la administración del Saccharomyces cerevisiae (SC), al inicio de la lactancia sobre el comportamiento reproductivo de vacas Holstein (H) y Carora (C) en la zona alta del estado Mérida, Venezuela. Cuarenta y un vacas H y 15 vacas C se dividieron en dos tratamientos: 1) grupo experimental (GE) 10 g/día de SC y 2) grupo control (GC) que no recibió SC, durante 105 días postparto (DPP). La alimentación basal era el pastoreo, más concentrado al momento del ordeño. Se evaluó el intervalo parto-primer cuerpo lúteo (IP1CL), intervalo parto-primer estro (IP1E) e intervalo parto-concepción (IPC). Los datos fueron analizados a través del análisis de varianza con la prueba de Tukey para la comparación de medias. No hubo diferencias estadísticas (P>0,05) entre tratamientos en las variables reproductivas en las vacas C. Sin embargo, en las vacas H del GE, el IP1CL fue menor (P<0,05) que en el GC (28,2 vs. 33,7 días). No obstante, el 100% de las vacas H y C del GE tuvieron su primer cuerpo lúteo antes de los 70 DPP. Igualmente, el 59,1% y el 75% de las vacas H y C tratadas expresaron su primer estro postparto antes de los 70 DPP. Los resultados indican que el uso estratégico del SC al inicio de la lactancia mejora el comportamiento reproductivo, posiblemente por la acción del SC en el rumen, incrementando la tasa de degradación de la fibra y la proporción de propionato.
Palabras clave: vacas lecheras postparto, levaduras, comportamiento reproductivo, cuerpo lúteo.
Effect of supplementation with Saccharomyces cerevisiae on reproductive performance on dairy herds in the mountain area of Mérida state
Summary
The aim of this study was to determine the strategic use of Saccharomyces cerevisiae (SC) from onset of lactation until 105 days postpartum and its effect on reproductive performance, on Holstein (H) and Carora (C) cows located in the mountain area of Merida state, Venezuela. Forty one H and 15 C cows were assigned to two treatments 1) experimental group (EG) fed with 10 g/day of SC, and 2) Control group (CG) that did not receive SC. C cows grazed pastures of Panicum maximum while H cows had access to pastures of Pennisetum clandestinum. All of the cows were fed at milking with a concentrate mixture (20% CP and 58% NDT) at a ratio of 1 kg:3 kg of milk. Reproductive traits such as interval calving-first corpus luteum (IP1CL), interval calving to first estrus (IP1E) and interval calving to conception (IPC) were evaluated. Data were analyzed by ANOVA and mean comparisons tested for significance using Tukey´s procedures. There were not statistical differences (P>0.05) among the reproductive variables between treatments and for the C cows. However, H cows of EG had shorter IP1CL compared to CG cows (28.2 vs 33.7 d). Moreover, 100% of the cows of the EG had their first CL prior to 70 days post calving. Similarly, 59.1% and 75% of the treated H and C cows, respectively, did show behavioral estrus before 70 days after parturition. Results from this study indicated that the strategic use of SC during the first 105 days post calving improved reproductive performance in dairy cows, perhaps by stimulating the action of SC in the rumen and the availability of nutrients in the mammary gland.
Key words: dairy cows, yeast culture, performance reproductive, corpus luteum.
INTRODUCCIÓN
La zona alta del estado Mérida presenta el sistema de producción de leche intensivo denominado ganadería de altura, constituido por la explotación de rebaños de vacas de alta producción de leche. Este sistema se caracteriza por programas de alimentación con un uso intensivo de los forrajes a pastoreo y altas cantidades de alimento concentrado, sin considerar el balance de la dieta. Esto trae como consecuencia pérdida de peso, condición corporal, bajo rendimiento en producción de leche al inicio de la lactancia y un deficiente comportamiento reproductivo postparto.
Los probióticos son aditivos no nutritivos, los cuales contienen diferentes preparaciones de levaduras (muertas, de panificación y los cultivos de levaduras) con efectos diversos sobre la estimulación de la flora ruminal., aportando enzimas, vitaminas y aminoácidos, los cuales tiene un efecto positivo sobre la respuesta productiva de los rumiantes (Dawson, 1993). Los beneficios pueden surgir de los metabolitos per se o por su interacción con otros microorganismos ruminales, mejorando el aprovechamiento de las fuentes nitrogenadas, tales como amonio y proteínas por parte de los microorganismos ruminales (Wholt et al., 1998). Dawson y Girard (1997) y Wholt et al. (1998) han demostrado resultados positivos con el uso de 10 g/vaca/día de Saccharomyces cerevisiae, dosis que garantiza un mínimo de 108 unidades formadoras de colonias (UFC/g); capaz de mantener de 5 a 10 x108 UFC en el rumen con 50 L de fluido, lo que representa por lo menos 4 x 108 UFC/ml de fluido ruminal.
Los cultivos de levaduras, específicamente, el S. cerevisiae (SC), actúan estimulando el crecimiento bacteriano, principalmente de las bacterias celulolíticas y las que utilizan el ácido láctico como substrato, mejorando las condiciones del ecosistema ruminal con beneficios sobre la nutrición animal, favoreciendo una pronta recuperación del balance energético negativo propio del inicio de la lactancia (Wallace y Newbold, 1993).
El efecto del balance negativo de energía, propio de las primeras semanas postparto, afecta la secreción de LH y disminuye la respuesta del ovario al efecto de LH, pudiendo ocurrir fallas en el desarrollo y la ovulación del primer folículo dominante y por ende la formación de tejido luteinizado (Hanzen, 1986; Schillo, 1992; Butler, 2000). Este tejido luteal es el responsable del incremento en los niveles de P4 en plasma, que indican el reinicio de la actividad ovárica postparto (Ramírez, 1995). Mejoras en el estado nutricional que repercutan sobre el balance energético conllevan a mayor eficiencia en la producción de carne y leche (Wallace y Newbold, 1993).
El objetivo general de la presente investigación fue determinar el efecto del uso estratégico del SC, al inicio de la lactancia, sobre el comportamiento reproductivo en vacas de alta producción de leche.
MATERIALES Y MÉTODOS
Se utilizaron 56 vacas lecheras de las razas H y C, ubicadas en dos rebaños. El Cuadro 1 presenta la descripción agroclimática de las unidades de producción. Los animales se distribuyeron al azar en dos grupos de tratamientos (Cuadro 2) para cada rebaño: 1) grupo experimental (GE), que recibieron 10g/día de SC, y 2) grupo control (GC).
|
Cuadro 1. Descripción agroclimática de las unidades de producción | ||
|
| ||
| Fundo | 1 | 2 |
|
| ||
| Grupo Racial | Carora | Holstein |
|
| ||
| Clima | Bosque seco premontano | Bosque muy húmedo montano |
| Altitud (msnm) | 1.050 | 2.800 |
| Temperatura (ºC) | 24 | 15 |
|
Cuadro 2. Distribución de las vacas por tratamiento en cada rebaño | |||
|
| |||
| Tratamiento | Rebaño | Total | |
|
| |||
| Carora | Holstein | ||
|
| |||
| -----------------------n---------------------- | |||
|
GE |
8 | 22 | 30 |
| GC | 7 | 19 | 26 |
|
| |||
| Total | 15 | 41 | 56 |
La dieta estuvo basada en el pastoreo de Pennisetum clandestinum y Panicum maximum en las vacas H y C respectivamente. Al momento del ordeño se suministró una ración de alimento concentrado en una relación de 1 kg de concentrado por cada 3 kg de leche producida, así como, una mezcla mineral completa, a razón de 100 g/d por animal.
La composición nutricional se determinó mediante el método de análisis proximal de Weende (Maynard et al., 1979) y para la obtención de fibra detergente neutra y ácida en el pasto, se utilizó el método de Van Soest (Maynard et al., 1979). En el cálculo de nutrientes digestibles totales (NDT) de los pastos, se utilizaron los valores de digestibilidad reportados por Bernal (1994). La metodología empleada en el cálculo del NDT, energía digestible (ED), energía metabolizable (EM) y energía neta de lactancia (ENL) se basó en la reportada por la NRC (1989). El Cuadro 3 resume la composición nutricional del pasto y alimento concentrado consumido por los animales.
|
Cuadro 3. Análisis proximal de la dieta | |||||||||
|
| |||||||||
| Fondo | Muestra | MS | PC | EE | FC | CEN | Ca | FDN | FDA |
|
| |||||||||
|
% | |||||||||
| 1 | Pasto | 25,0 | 8,30 | 1,49 | 34,28 | 14,77 | 0,54 | 66,16 | 41,95 |
| Concentrado | 87,8 | 19,98 | 1,01 | 5,87 | 8,14 | 1,21 | |||
| 2 | Pasto | 21,0 | 12,49 | 1,18 | 27,64 | 15,81 | 0,38 | 64,69 | 34,16 |
| Concentrado | 88,37 | 21,44 | 0,67 | 6,65 | 8,92 | 1,26 | |||
|
| |||||||||
|
MS: materia seca; PC: proteína cruda; EE: extracto etéreo; FC: fibra cruda; CEN: cenizas; Ca: calcio; FDN: Fibra detergente neutra y FDA: Fibra detergente ácida. | |||||||||
Se estudiaron las variables: Intervalo parto-primer cuerpo lúteo (IP1CL), intervalo parto-primer estro (IP1E) e intervalo parto-concepción (IPC). Posterior al parto y semanalmente las vacas se palparon hasta la determinación del primer cuerpo lúteo.
En cada rebaño se aplicó un diseño experimental completamente aleatorizado, los datos fueron analizados a través del análisis de variancia de Mínimos Cuadrados, considerando como efectos principales el tratamiento y número de partos (NP). Se utilizó la prueba de Tukey para la comparación de las medias. El modelo lineal utilizado en el análisis de los datos fue el siguiente: Yij = µ + Ti + NPj +Eij, donde: Yij: la n-ésima vaca en el i-ésimo tratamiento en el j-ésimo número de partos; µ: media general; Ti: i-ésimo tratamiento; NPj: j-ésimo número de partos; y Eij: error experimental de la n-ésima vaca en el i-ésimo tratamiento en el j-ésimo número de partos. La interacción T por NP no se incluyo en el modelo final por determinarse en análisis previos no significancia.
RESULTADOS Y DISCUSIÓN
El IP1CL fue afectado por la inclusión del SC, siendo de menor duración en el GE de las vacas H (28,2 ± 1,7 días; P<0,05), las cuales presentaron su primer cuerpo lúteo 5 días antes que el GC (33,7 ± 3,4 días) (Figura 1). Igualmente, el 100% de las vacas del GE presentó su primer cuerpo lúteo antes de los 70 DPP vs. 94,8% de las vacas del GC.
Por otra parte, la incorporación de SC no afectó el IP1CL en las vacas C, siendo su duración de 37,2 ± 9,1 días en el GC y de 37,9 ± 5,8 días en el GE. A pesar de no haber diferencias estadísticas, el 100% de las vacas del GE presentó su primer cuerpo lúteo antes de los 70 DPP vs. 85,7% del GC.
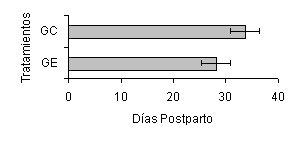 |
| Figura 1. Efecto del Saccharomyces cerevisae, sobre la duración del intervalo parto-primer cuerpo lúteo en vacas Holstein. |
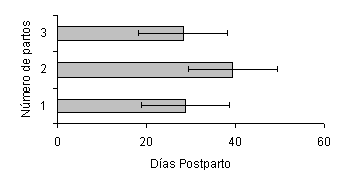
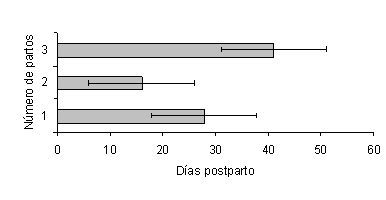
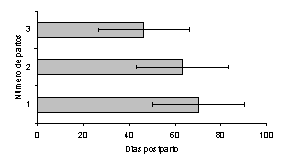
El NP afectó la duración del IP1CL (Figura 2), siendo menor en las vacas H de primer parto (28,8± 3,3 días; P<0,01) y de 3 ó más partos (28,2 ± 1,7 días; P<0,01), las cuales presentaron su primer cuerpo lúteo 10 días antes que las vacas H de 2 partos (39,5 ± 5,8 días). Igualmente, en las vacas C el NP afectó la duración del IP1CL (Figura 3), siendo menor en las vacas de 2 partos (16 días; P<0,01) y de primer parto (27,8 ± 8 días; P<0,01); las cuales presentaron su primer cuerpo lúteo 25 y 14 días antes, respectivamente, que las vacas de 3 ó más partos (41 ± 5,8 días).
 |
|
Figura 2. Efecto del número de partos, sobre la duración del intervalo parto-primer cuerpo lúteo en vacas Holstein. |
 |
|
Figura 3. Efecto del número de partos, sobre la duración del intervalo parto-primer cuerpo lúteo en vacas Carora. |
El IP1CL es un factor indicativo de la reanudación de la actividad ovárica. Según lo señalado por Hanzen (1986), Schillo (1992) y Butler (2000), el efecto del balance negativo de energía propio de las primeras semanas postparto, afecta la secreción de LH y disminuye la respuesta del ovario a la estimulación por LH. Esto puede causar fallas en la ovulación del primer folículo dominante y por ende la formación de tejido luteinizado, responsable del incremento de los niveles de P4 en plasma, indicador del reinicio de la actividad ovárica postparto (Ramírez, 1995).
Los resultados obtenidos coinciden con los reportados por Párraga (1988) en vacas Holstein adultas estabuladas en ambiente bosque seco tropical y son inferiores a los reportados por Bastidas (1982) en animales Bos indicus, a pastoreo sin suplementación, ubicados en la misma zona de vida y a los señalados por Hidalgo (1993) en vacas mestizas Holstein y Pardo Suizo a pastoreo y suplementadas con bloques multinutricionales y concentrado y localizadas en el ambiente de bosque seco tropical.
Los valores obtenidos para el IP1CL en las vacas H permiten inferir que el uso estratégico del SC durante un período crítico como es el inicio de la lactancia favoreció el reinicio de la actividad ovárica postparto en las vacas suplementadas con el SC. En el caso de las vacas C, la adición del SC no redujo el IP1CL. Sin embargo, se pueden reportar los beneficios de su administración al observar que el 100% de las vacas C del GE presentaron su primer cuerpo lúteo antes de los 70 DPP. Esta respuesta pudiese ser explicada por el efecto positivo que ejerce el SC sobre el rumen, mejorando: i) tasa de digestibilidad de los componentes de la dieta, ii) porcentaje de degradabilidad del forraje, iii) cambios en el patrón de fermentación ruminal, iv) modificación del pH ruminal, v) cambios en el número de microorganismos del rumen y vi) interacción bacterias dieta (Arambel y Kent, 1990; Wohlt et al., 1991; Wohlt et al., 1998) e incrementando la disponibilidad de energía (Robinson, 1997), en especial cuando se ofrecen altas cantidades de concentrado.
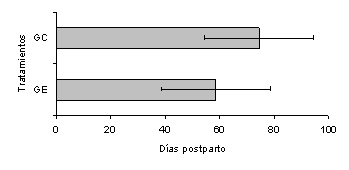
La administración de SC no afectó la duración del IP1E en las vacas H, siendo de 51,8 ± 3,4 días para el GC y de 56,4 ± 5,1 días para el GE. En las vacas C, no hubo efecto del SC sobre la duración del IP1E, siendo de 74,5 ± 5,5 días en el GC y 58,6 ± 11,4 días en el GE (Figura 4). A pesar de no haber diferencias estadísticas, se observa que las vacas C del GE manifestaron su primer celo postparto 16 días antes que las vacas del GC. El 75% de las vacas C del GE expresó su primer estro antes de los 70 DPP vs. 42,8% de las vacas del GC.
 |
|
Figura 4. Efecto del Saccharomyces cerevisiae sobre la duración del intervalo parto-primer estro lúteo en vacas Carora. |
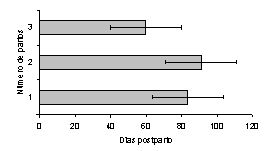
El NP afectó la duración del IP1E (Figura 5) en las vacas H, siendo menor en las vacas de 3 ó más partos (46,4 ± 4,4 días; P<0,01), las cuales expresaron su primer estro postparto 24 días antes que las vacas de primer parto (70,2 ± 11,7 días) y 17 días antes que las vacas de 2 partos (63,3 ± 8,2 días). Igual respuesta se observó en las vacas C, en las cuales el NP afectó la duración del IP1E (Figura 6), siendo menor en las vacas de 3 ó más partos (59,8 ± 8,1 días; P<0,05), las cuales expresaron su primer estro postparto 24 días antes que las vacas de primer parto (83,5 ± 8,5 días) y 32 días antes que las vacas de 2 partos (91 días).
 |
|
Figura 5. Efecto del Número de partos, sobre la duración del intervalo parto-primer estro lúteo en vacas Holstein (P<0,01). |
 |
|
Figura 6. Efecto del Número de partos, sobre la duración del intervalo parto-primer estro lúteo en vacas Carora (P<0,05). |
Los resultados obtenidos para el IP1E en las vacas H y C suplementadas con levaduras fueron inferiores a los reportados por González-Stagnaro (1992), en vacas mestizas doble propósito sin suplementación y coinciden con lo reportado por el mismo autor en vacas Pardo Suizo y H estabuladas y en vacas mestizas a pastoreo suplementadas con concentrado (Ramírez, 1995). Por otra parte son menores a los reportados por Vásquez (1999) en vacas Brahman, suplementadas con proteína sobrepasante.
Por otra parte, los resultados obtenidos para el IP1E, coinciden con los mencionados por Bastidas et al. (1998), en vacas mestizas doble propósito y Carora con puerperio normal, con un manejo alimenticio a pastoreo y concentrado al momento del ordeño y con los valores para el IP1E reportado por Soto et al. (1998), en vacas mestizas doble propósito expuestas a toros adultos a partir de los 15 DPP.
La implementación de estrategias de suplementación al inicio de la lactancia, mejora el IP1E en las vacas lecheras (Butler, 2000), destacando la importancia del plano nutricional sobre la respuesta reproductiva de los animales al inicio de la lactancia (Hanzen, 1986; Schillo, 1992
Los valores de IP1E obtenidos en las vacas H de primer parto coinciden con lo reportado por Ramírez (1995) y difieren con relación a las vacas de 2 partos. No obstante, los valores obtenidos en las vacas C de 1 y 2 partos, son superiores a los reportados por dicho autor en vacas con igual número de partos.
La adicción de SC a la dieta de las vacas H no afectó la duración del IPC, siendo de 80 ± 4 días en el GC y de 76,8 ± 6,7 días en el GE, teniéndose que el 18,2% de las vacas de este grupo concibió antes de los 70 DPP vs. 10,5% del GC. El 50% de las vacas H del GE quedó gestante al primer servicio postparto, mientras que en el GC sucedió solo en el 40% de las vacas. Igual respuesta se obtuvo en las vacas C, grupo en el cual el IPC no fue afectado por la administración del SC siendo el IPC en las vacas del GC de 79 ± 9,1 días y de 79,2 ± 7,5 días en las vacas del GE, observándose que el 50% de las vacas de este grupo concibió antes de los 70 DPP vs. 57,2% del GC. El porcentaje de vacas C gestantes al primer servicio en el GC fue de 83%, mientras que en el GE fue del 50%.
Por otra parte, el NP, mostró una tendencia (P<0,10) sobre el IPC en las vacas H y C, siendo menor en las vacas con 3 ó más partos las cuales concibieron 17 y 15 días antes que las vacas de primer parto y 6 días antes que las vacas de 2 partos, respectivamente.
Los resultados para el IPC obtenidos en el presente estudio fueron inferiores a los reportados por González-Stagnaro (1992) en vacas doble propósito ordeñadas con becerro y suplementadas con concentrado. Así mismo son menores a los reportados por Ramírez (1995) en vacas mestizas H y Pardo Suizo a pastoreo y suplementadas con concentrado y a los obtenidos por Bastidas et al. (1998) en vacas mestizas H y Carora a pastoreo mas concentrado, con puerperio normal.
La duración del IPC varía en relación al momento en que las vacas ingresen al período de servicio, la tasa de detección de celo y la fertilidad (González-Stagnaro, 1992). En el presente trabajo, se obtuvo una reducción del 18% en la duración del intervalo entre partos en las vacas H y del 5% en las vacas C. Estos resultados coinciden con Didley (1990), quien menciona una mejora del 4% en el intervalo entre partos con el suministro de levaduras, sin que esta mejora sea estadísticamente significativa.
El uso del SC al inicio de la lactancia promovió el reinicio de actividad luteal durante el primer mes postparto, independientemente del NP; y bajo condiciones de manejo adecuadas, esto favorece la expresión del primer celo postparto, evento que indica las condiciones de funcionamiento endocrino favorables para reiniciar la actividad reproductiva postparto de la hembra bovina. En consecuencia, la aplicación de estrategias que mejoren el plano nutricional durante el período crítico del inicio de la lactancia, ejercen efectos positivos sobre el comportamiento reproductivo, permitiendo un ingreso más temprano al período de servicios y por ende garantizando intervalos entre partos menores a 13 meses.
CONCLUSIONES
El presente estudio ofrece una evaluación inicial del efecto de las levaduras sobre el sistema de producción de leche de la zona alta del estado Mérida, Venezuela. El uso estratégico del SC, durante los primeros 105 días del postparto favorece un pronto reinicio de la actividad ovárica postparto en las vacas Holstein, aspecto importante, ya que permite un pronto ingreso de las vacas al período de servicio y una obvia reducción del período de espera voluntario.
LITERATURA CITADA
Bastidas P. 1982. Comportamiento reproductivo postparto en ganado Brahman. Trabajo de ascenso. Universidad Central de Venezuela, Facultad de Ciencias Veterinarias, Maracay, Venezuela.
Bastidas P., N. Guerrero, M. Manzo y T. Díaz. 1998. Manejo reproductivo postparto de vacas lecheras. En González-Stagnaro C., N. Madrid-Bury y E. Soto-Belloso (Eds.) Mejora de la Ganadería de Doble Propósito. Astro Data. Maracaibo. pp. 413-426.
Butler W. R. 2000. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci., 60 61: 449457.
Bernal E. J. 1994. Pastos y forrajes, producción y manejo. 3ra Ed. Banco Ganadero. Colombia. p. 470.
Dawson K. A. 1993. The use of yeast culture in animals feeds: a scientific application of direct fed microbials and challenges of the future. En: T. P. Lyons (Ed.). Biotechnology in the Feed Industry, proceedings of Alltechs Ninth Annual Symposium. USA. pp. 169-172.
Dawson K. y Girard I. D. 1997. Biochemical and physiological basis for the stimulatory effects of yeast preparations on ruminal bacteria. En Lyons T.P. (Ed.) Biotechnology in the Feed Industry. Proc. Alltechs 9th Annual Symposium. USA. pp. 293-305.
Didley D. 1990. Effect of Yea Sacc on performance of cows at the Maddox dairies in California. Yea Sacc 1026 Technical Dossier. Alltech Inc. USA. p.32.
Hanzen Ch. 1986. Endocrine regulation of postpartum ovarian activity in cattle: a review. Reprod. Nutr. Develop., 26: 1219-1239.
Hidalgo T. M .E. 1993. Determinación de la actividad ovárica y comportamiento reproductivo en vacas doble propósito con diferente régimen de alimentación. Tesis de Grado. Universidad Central de Venezuela, Facultad de Agronomía. Maracay, Venezuela. p. 82.
Maynard L. A., J. K. Loosli, H. F. Hintz y R. G. Warner. 1979. Nutrición Animal. 4ta Ed.. McGraw-Hill. Mexico. pp. 94.
NRC (National Research Council). 1989. Nutrient Requirements of Dairy Cattle. 6ta Ed. National Academy Press, Washington.
Párraga I. C. 1988. Estudio de la actividad ovárica postparto a través de palpación rectal y cuantificación hormonal en vacas Holstein. Tesis de Maestría. Postgrado en Reproducción Animal y Tecnología de la Inseminación Artificial. Universidad Central de Venezuela, Facultad de Ciencias Veterinarias. Maracay. Venezuela. 81p.
Ramírez L. N. 1995. Factores que afectan el período vacío en vacas Carora y Mestizas. En Madrid-Bury N. y E. Soto Belloso (Eds.). Manejo de la Ganadería de Doble Propósito. Astro Data. Maracaibo. pp. 465-486.
Schillo K. K. 1992. Effects of dietary energy on control of luteinizing hormone secretion in cattle and sheep. J Anim. Sci., 70:1271-1272.
Soto E., G. Portillo y G. Soto. 1998. Avances en el manejo reproductivo de la vaca problema en ganadería de doble propósito. En González-Stagnaro C., N. Madrid-Bury y E. Soto-Belloso (Eds.). Mejora de la Ganadería de Doble Propósito. Astro Data. Maracaibo. pp. 427-442.
Vásquez B. 1999. Comportamiento reproductivo de vacas Brahman de primera lactancia suplementadas con proteína sobrepasante. Tesis de Maestría. Postgrado en Reproducción Animal y Tecnología de la Inseminación Artificial Universidad Central de Venezuela Facultad de Ciencias Veterinarias. Maracay, Venezuela. p. 60.
Wallace R. J. y C. J. Newbold. 1993. Rumen fermentation and its manipulation: the development of yeast cultures as feed additives. En Lyons T.P. (Ed.) Biotechnology in the Feed Industry. Proc. Alltechs 9no Annual Symposium. USA. pp. 173-192.
Wohlt J. E., T. T. Corcione y P. K. Zajac. 1998. Effect of yeast on feed intake and performance of cows fed diets based on corn silage during early lactation. J. Dairy Sci., 81: 1345-1352.